





基于适应性免疫的T细胞免疫疗法自出现之初便迅猛改变着肿瘤治疗的格局,其通过靶向和动员T细胞、恢复并增强T细胞的抗肿瘤活性从而有效消灭恶性细胞。根据作用机理,基于T细胞的癌症免疫疗法主要分为两类[1]:第一类针对如PD-1,PD-L1等免疫抑制分子。其代表性疗法为免疫检查点抑制剂。而这也是首批成功恢复T细胞抗肿瘤活性的免疫疗法之一,最早可追溯至2011年FDA批准使用伊匹木单抗(Ipilimumab,抗CTLA-4)来治疗转移性黑色素瘤[2]。但这种疗法仅对部分患者有效[3],因此专注于免疫刺激通路的第二类疗法便应运而生。其代表性疗法为嵌合抗原受体(CAR)T细胞和T细胞衔接器(T cell engager,TCE)。CAR-T细胞疗法具有高度特异性和个体化治疗等诸多优势,但其高昂的生产成本、相对较长的生产周期以及化疗后T淋巴细胞数量和功能的下降,限制了其广泛应用。而TCE作为一种新兴的癌症免疫疗法,聚焦了药物研发者的目光。本文将介绍TCE分子的研究进展及其药代动力学特征。
TCE分子及其设计与作用机理
抗肿瘤的双抗分子设计通常是使一个蛋白结构域与肿瘤相关抗原(TAA)结合(例如:BCMA, CD19, DLL3等[4]),而另一个结构域则与能够促进抗肿瘤效应的靶点相结合。抗肿瘤活性一般可以通过以下三种机制实现:
直接与免疫效应细胞的表面抗原结合,例如内源性T细胞表面的CD3,NK细胞表面的CD16,或巨噬细胞表面的CD47;
与调节T细胞相关反应的受体相结合;
与其它信号通路相互作用[5]。
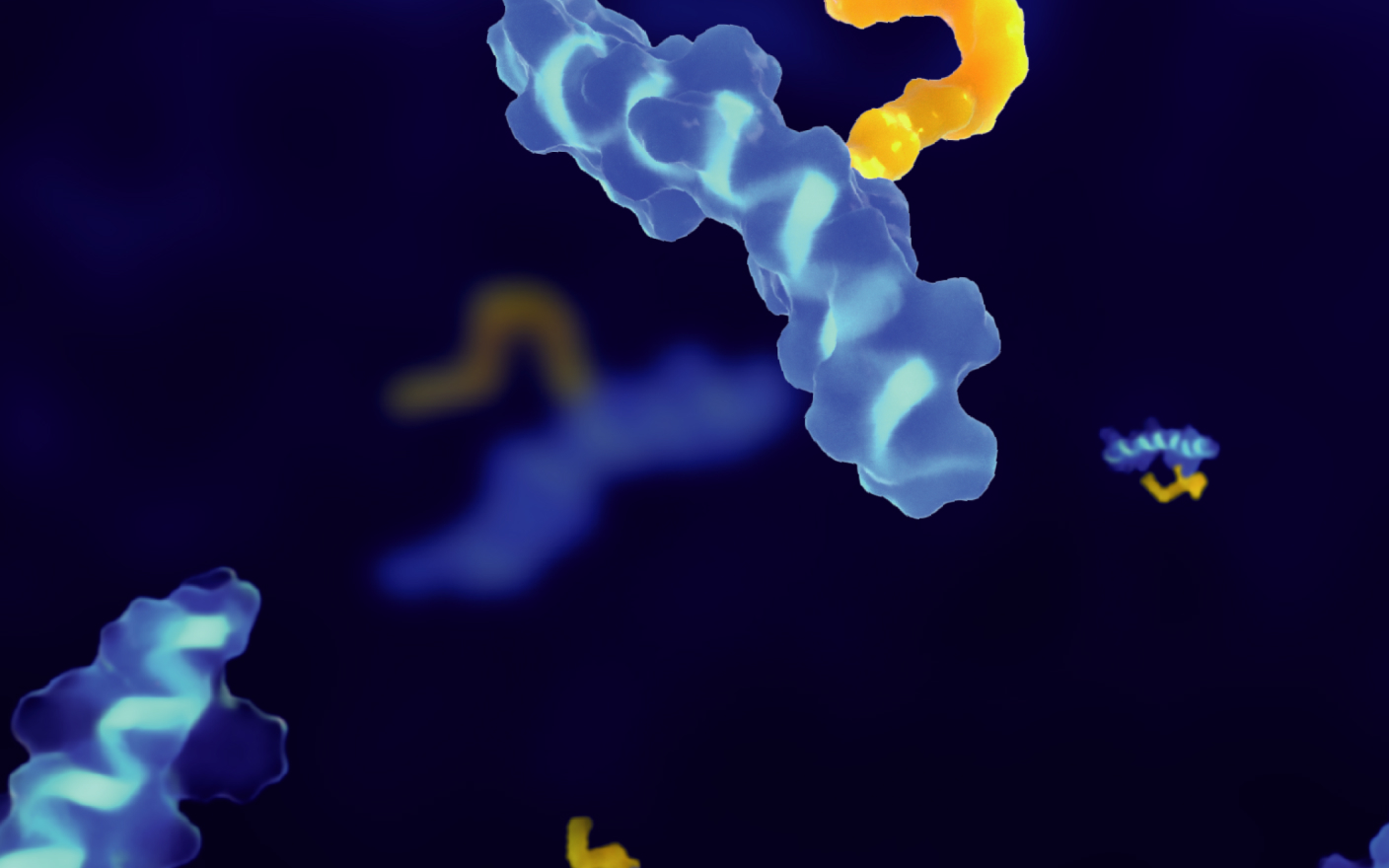
最为常见的TCE双抗分子设计则是采用了上述第一种方式,即同时靶向TAA与T细胞表面抗原。由于T细胞受体(TCR)中CD3链的恒定性,CD3通常被选择作为T细胞的表面靶点。通过同时结合TAA和CD3,TCE有效地实现了T细胞与癌细胞的直接互动,无需依赖主要组织相容性复合体(MHC)即可实现穿孔素和颗粒酶的释放,发挥强力的杀瘤功效。
基于结构和作用机理,TCE双抗分子可以分为两大类:即含有、或不含有Fc结构域的TCE分子。含有Fc结构域的TCE双抗分子,具有更长的半衰期与更高的稳定性。但与此同时,完整的Fc结构域会降低T细胞的迁移能力并限制其抗肿瘤活性[6]。此外,Fc结构域介导的抗体依赖细胞毒性(ADCC)和补体依赖细胞毒性(CDC)也可能导致TCE双抗在体内循环中引发不必要的非特异性免疫反应。而不含有Fc结构域的TCE分子则具有较小的分子尺寸。虽然其血浆半衰期较短,但具有更好的扩散性与组织穿透能力,可能降低临床的使用剂量。常见的不含Fc结构域的TCE分子包括纳米抗体,和由来自抗TAA和抗CD3单克隆抗体的两个单链可变片段(scFv)通过短链串联连接而成的BiTE分子[1]。

图1. BiTE分子的结构与作用机理[7]
TCE的适应症及其优势
与CAR-T细胞疗法相比,TCE双抗是一种小而美的替代疗法。除了可以免除CAR-T需进行的基因改造与体外细胞扩增之外,其诱导的肿瘤细胞杀伤效率也较高。首先,随时间推移,单个T细胞能够通过TCE与多个肿瘤细胞进行桥接,从而实现肿瘤细胞的连续杀伤。其次,TCE诱导的T细胞激活还会导致细胞因子的释放和T细胞的增殖,更进一步扩大了可用于TCE介导的肿瘤细胞杀伤的T细胞群体。此外,由于TCE介导的肿瘤细胞识别不依赖于MHC的作用,从而可以克服肿瘤细胞通过下调MHC I类分子的表达来逃避免疫监视的策略[3]。因此,TCE的高效性和灵活性使其成为当前癌症免疫治疗领域的重要研究方向之一。
目前,TCE药物的研发已从早期靶向B细胞相关抗原(如CD19,CD20等)的血液系统恶性肿瘤领域扩展至靶向TAA的实体瘤领域,并展现出相关治疗潜力。此外,基于对B细胞的深度清除能力,TCE药物在如类风湿性关节炎(RA)、系统性红斑狼疮(SLE)等B细胞介导的自身免疫性疾病领域也显示出广阔的应用前景。
已上市的TCE药物一览
表1. 全球已获批上市TCE药物列表[8]
药品名称(商品名) | 靶点 | 适应症 | 研发公司 | 技术平台 | 获批时间 |
卡妥索单抗 Catumaxomab (REMOVAB) | EpCAM×CD3 | 恶性胸腹水 | Neovii Biotech GmbH凌腾医药 | Triomab | 2009.05 EMA* |
贝林妥欧单抗 Blinatumomab (BLINCYTO) | CD19×CD3 | 前体B细胞淋巴细胞白血病、B细胞急性淋巴细胞白血病 | Amgen 百济神州 | BiTE | 2014.12 FDA |
Tebentafusp (KIMMTRAK) | Gp100×CD3 | 葡萄膜黑色素瘤 | Immunocore | - | 2022.01 FDA |
特立妥单抗 Teclistamab (TECVAYLI) | BCMA×CD3 | 多发性骨髓瘤 | Genmab Janssen | Dubody | 2022.8 EMA |
莫妥珠单抗 Mosunetuzumab (LUNSUMIO) | CD20×CD3 | 滤泡性淋巴瘤 | Roche | CrossMab/KIH | 2022.06 EMA 2024.12 NMPA |
格罗菲妥单抗 Glofitamab (COLUMVI) | CD20×CD3 | 弥漫性大B细胞淋巴瘤 | Roche | CrossMab/KIH | 2023.06 FDA |
塔奎妥单抗 Talquetamab (TALVEY) | GPRC5D×CD3 | 多发性骨髓瘤 | Janssen | Dubody | 2023.08 FDA 2025.02 NMPA |
埃纳妥单抗 Elranatamab (ELREXFIO) | BCMA×CD3 | 多发性骨髓瘤 | Pfizer | Dubody | 2023.08 FDA 2025.03 NMPA |
塔拉妥单抗 Tarlatamab (MDELLTRA) | DLL3×CD3 | 小细胞肺癌 | Amgen | HLE BiTE | 2024.05 FDA |
奥尼妥单抗 Odronextamab (ORDSPONO) | CD20×CD3 | 滤泡性淋巴瘤,弥漫性大B细胞淋巴瘤 | Regeneron | Veloci-Bi | 2024.08 EMA |
* 2017年退市
根据已批准上市的TCE药物列表,目前多数的TCE药物都用于治疗血液系统恶性肿瘤。其靶点包括CD19,CD20,BCMA和GPRC5D;适应症涵盖前体B淋巴细胞白血病,B细胞急性淋巴白血病,滤泡型淋巴瘤,弥漫性大B细胞淋巴瘤,多发性骨髓瘤等。
TCE目前主要应用于血液系统肿瘤的原因之一是,正常细胞中也表达这类抗原,其所导致的可能的非肿瘤靶向毒性,在血液瘤领域相比实体瘤而言更为可控。举例来说,靶向B细胞表面的CD19可能会导致B细胞再生障碍和低丙种球蛋白血症,并极大地增加感染风险;但这些感染可以通过临床警戒和及时的抗生素使用得以控制[5]。
TCE药物的研究挑战与进展
除了上文介绍的缘由之外,在实体瘤领域,TCE药物的开发还面临着诸多挑战,包括频发的细胞因子释放综合征(CRS)、复杂的肿瘤微环境(TME)、有限的肿瘤内T细胞可及性,以及肿瘤抗原缺失和缺乏肿瘤抗原特异性等[7,9]。但由于实体瘤类型占据了癌症种类的90%以上,因此在该领域,TCE药物仍在火热研发中。
目前,旨在治疗实体瘤的TCE疗法进展包括但不限于:
通过降低CD3的亲和力和/或改变TAA的数量与亲和力来减少细胞因子释放综合征的发生[9];
通过靶向胞内肿瘤新抗原或设计条件性激活的TCE分子来克服非肿瘤靶向毒性[10,11];
通过增加靶向TAA的数量(设计三抗分子)来提高抗肿瘤活性,同时避免肿瘤抗原缺失的干扰[12];
通过TCE mRNA的原位表达来实现TCE分子在肿瘤部位的富集[13];
通过与抗PD-1/PD-L1单抗的联合治疗或设计靶向PD-L1的TCE分子来提升TCE的杀瘤效力、避免肿瘤细胞的免疫逃逸;
通过与腺苷受体拮抗剂或IL-10抑制剂的联合治疗来破坏免疫抑制的肿瘤微环境[9]。
现今,多家药企也已积极布局TCE三抗药物管线,例如Novartis推出了靶向CD19×CD2×CD3的三抗产品PIT565,而GSK也引进了靶向CD19×CD20×CD3的国产三抗分子CMG1A46。近期,AstraZeneca拟用于治疗B型非霍奇金淋巴瘤、系统性红斑狼疮的三抗分子AZD5492(CD20×TCR×CD8)也在中国数度获批临床。
TCE药物的药代动力学检测考量
PK检测
截至目前,监管机构尚未发布针对TCE药物的个药指南,因此针对TCE药物的药代动力学研究可以参考FDA于2019年4月发布的《Bispecific Antibody Development Programs》指导草案,和NMPA于2022年11月发布的《双特异性抗体抗肿瘤药物临床研发技术指导原则》。
TCE药物的PK和ADA实验设计和生物分析可参考已发布文章:双特异性抗体药物盘点及其药代动力学特征。
细胞因子检测
除常规双抗药物的PK和ADA检测外,TCE药物通常会引发严重的细胞因子释放综合征(CRS)以及免疫效应细胞相关神经毒性综合征(ICANS),且这两者的发生通常相互伴随。因此,在TCE药物的临床前研究中,细胞因子的检测显得尤为重要[14]。TCE药物的细胞因子考量主要集中在各类辅助型T细胞(helper T cell, Th),如:Th1、Th2和Th17。同时,根据药物的具体靶点,可能还需增加其他细胞因子或趋化因子的检测。例如在CD3×CD19双抗Blinatumomab的细胞因子检测中包含了IL-2、IL-4、IL-6、IL-8、IL-10、IL-12、TNF-α和IFN-γ;其中IL-6、IL-10和IFN-γ的升高尤为显著[15]。
此外,由于细胞因子的释放通常是一过性的,因此采样点的设计应更加密集。建议在给药前,以及首次和末次(如有)给药后的数小时内及24小时分别进行细胞因子的测定,尤其是针对与药物作用机制相关靶点所产生的细胞因子。
目前,药明康德DMPK对细胞因子的检测包括基于流式细胞术的CBA方法(cytometric bead array)以及基于电化学发光的MSD方法。这两种方法均适用于多因子检测,能够显著节省所需样本。其中,MSD平台能够检测到fg/mL级别的样本,弥补了流式CBA在灵敏度方面的不足。
免疫分型检测
为了更好地建立TCE药物的PK/PD模型,检测T细胞(及其他主要免疫细胞群)的数量、T细胞的活化/增殖状态、以及靶细胞的数目是有必要的。在Blinatumomab的临床前和临床研究中均发现在给药后,外周血中的T细胞通常会黏附于血管内皮或迁移至组织,从而导致T细胞数量的瞬时减少。而T细胞数目的检测通常也会伴随其他主要免疫细胞群的监测,如B细胞、自然杀伤细胞(natural killer cells, NK cells)以及单核细胞(monocytes),以评估药物介导的细胞裂解。
另一方面,T细胞在TCE药物的作用下会被显著激活,因此对于T细胞活化状态的监控也是有必要的,通常选用的指标包括CD25和/或CD69,有时还需同步检测该细胞群的增殖情况[16]。例如在Blinatumomab的研究中, B细胞数目的监控以及靶细胞数目的检测常作为重要的PD指标[15]。此类检测依赖于流式细胞术的免疫分型技术。
目前,药明康德DMPK对于全血,骨髓以及各类组织的免疫分型检测积累了丰富的经验。基于细胞表面、胞内或核内不同指标的表达差异,以及细胞大小和颗粒度的异质性,我们能够通过不同荧光标记的抗体或其他标记物精准区分与统计不同细胞类型的比例(以及液体组织中的绝对数目)。针对TCE药物,我们已在全血中为常见的T细胞、B细胞、NK细胞以及单核细胞建立免疫分型检测的染色面板。通过动态监测免疫细胞的变化情况,更深入地评估药物对各类细胞的影响,从而为该类药物的临床前研究提供更多信息。
案例分享
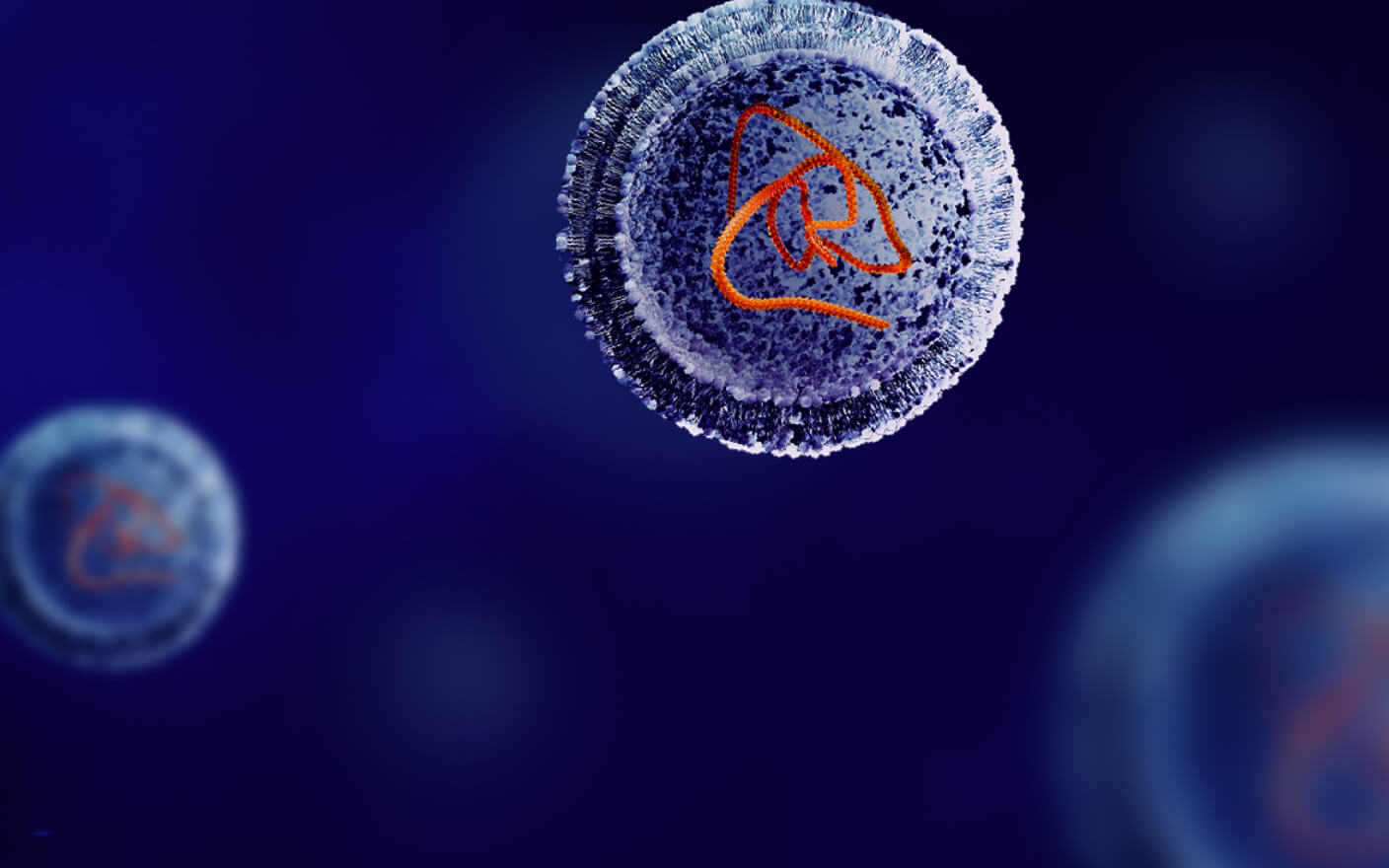
如前文所述,设计条件性激活的TCE分子(probody)以克服非肿瘤靶向毒性是当前优化TCE分子设计的热点方向之一。2019年,Seagen公司在Nature Biotechnology上发表了一项创新的通用型抗体掩蔽策略[17],通过将具有高亲和力的平行异源二聚体螺旋结构域与抗体N端融合,从而实现对抗体的结合能力和生物学功能的可逆性抑制。此外,该设计利用肿瘤微环境中高表达的基质金属蛋白酶(如MMP-2和MMP-9)对遮蔽肽进行特异性切割,通过选择性恢复抗体活性,大幅降低了脱靶毒性的发生风险。

图2. Probody示意图[17]

图3. A掩蔽与未掩蔽的抗小鼠CD3抗体和表达小鼠CD3的HT-2细胞的结合能力;B掩蔽与未掩蔽的抗小鼠CD3抗体的药-时曲线[17]
基于抗CD3抗体的已知靶向毒性,研究者们使用抗小鼠CD3抗体验证了螺旋遮蔽肽是否可以有效阻止抗体与淋巴细胞表面的CD3蛋白相结合。如图3A所示,在高达2 μM的浓度下,遮蔽肽成功“掩护”了抗小鼠CD3抗体,使其未与CD3+ HT-2细胞发生结合。
此外,在对BALB/c小鼠(n=3)进行静脉注射给药0.5 mg/kg的抗小鼠CD3抗体后,未掩蔽的抗体由于CD3广泛表达所致的抗原沉默效应(antigen sink effect),在给药2天后即无法在小鼠血浆中检测到。相比之下,掩蔽后的抗小鼠CD3抗体则在15天内表现出优异的循环稳定性(图3B)。

图4. 小鼠给药掩蔽或未掩蔽的抗小鼠CD3抗体后IL-2和IFN-γ的水平变化[17]
同时,掩蔽策略亦能够有效避免促炎细胞因子的过度释放。如图4所示,未掩蔽的亲本抗体在给药后4小时显著提高了动物体内细胞因子IL-2和IFN γ的浓度,而连接遮蔽肽的抗小鼠CD3抗体则很大程度地削弱了相关靶向毒性。
这种基于螺旋肽的抗体掩蔽策略凭借其通用型设计、高效的掩蔽能力、选择性激活抗体活性、以及显著优化的药代动力学特征,拓宽了TCE药物的治疗窗,且有助于临床给药剂量的爬升。这种设计为靶向在正常组织中表达而无法通过传统方法触及的抗原提供了新的可能性,并有望加速TCE药物的开发与临床转化进程。
结语
TCE药物凭借其高效激活T细胞、精准杀伤肿瘤细胞的能力,已成为癌症治疗领域的一颗亮眼明星。目前,多种TCE双抗在血液瘤的治疗中已展现出相关疗效;与此同时,通过优化分子结构、探索多靶点策略、联合免疫检查点抑制剂等方式,TCE药物在实体瘤治疗中的应用潜力也逐渐崭露头角。未来,随着更多临床数据的积累和技术创新的推进,TCE药物有望为更多癌症患者带来新的福音。
药明康德DMPK在TCE药物研究领域积累了丰富的经验。目前已针对TCE药物的PK分析、ADA免疫原性、细胞因子、免疫分型及临床生化检测建立了整合型生物分析平台,为客户提供一体化服务,助力新药研发进程。
作者:王骁琦,颜欢,潘杰,程起干,金晶
编辑:钱卉娟,富罗娜·克里木
设计:倪德伟,张莹莹
药明康德DMPK依托中国(上海、苏州、南京和南通)和美国(新泽西)的研发中心,提供从早期筛选、临床前开发、到临床研究阶段的综合型药代动力学服务,助力您快速推进药物研发流程。拥有上千人的研发团队,服务超1600家全球客户,具有超过十五年的新药申报经验,已成功支持超过1700个新药临床研究申请(IND)。
点击此处与我们的专家进行联系。
参考
[1] Zhou S, Liu M, Ren F, et al. The landscape of bispecific T cell engager in cancer treatment[J]. Biomarker research, 2021, 9(1): 38.
[2] Wei S C, Duffy C R, Allison J P. Fundamental mechanisms of immune checkpoint blockade therapy[J]. Cancer discovery, 2018, 8(9): 1069-1086.
[3] Arvedson T, Bailis J M, Britten C D, et al. Targeting solid tumors with bispecific T cell engager immune therapy[J]. Annual Review of Cancer Biology, 2022, 6(1): 17-34.
[4] Einsele H, Borghaei H, Orlowski R Z, et al. The BiTE (bispecific T‐cell engager) platform: development and future potential of a targeted immuno‐oncology therapy across tumor types[J]. Cancer, 2020, 126(14): 3192-3201.
[5] Shanshal M, Caimi P F, Adjei A A, et al. T-Cell engagers in solid cancers—Current landscape and future directions[J]. Cancers, 2023, 15(10): 2824.
[6] Wang L, Hoseini S S, Xu H, et al. Silencing Fc domains in T cell–engaging bispecific antibodies improves T-cell trafficking and antitumor potency[J]. Cancer immunology research, 2019, 7(12): 2013-2024.
[7] Goebeler M E, Bargou R C. T cell-engaging therapies—BiTEs and beyond[J]. Nature Reviews Clinical Oncology, 2020, 17(7): 418-434.
[8] 海通国际,TCE双抗行业研究
[9] Singh A, Dees S, Grewal I S. Overcoming the challenges associated with CD3+ T-cell redirection in cancer[J]. British journal of cancer, 2021, 124(6): 1037-1048.
[10] Zhang J, Guo Q, Wang Q, et al. CD3‐T‐cell‐engager (TCE) therapies to overcome solid tumors: Beyond BiTEs[J]. MedComm–Biomaterials and Applications, 2022, 1(2): e20.
[11] Trang V H, Zhang X, Yumul R C, et al. A coiled-coil masking domain for selective activation of therapeutic antibodies[J]. Nature biotechnology, 2019, 37(7): 761-765.
[12] Seung E, Xing Z, Wu L, et al. A trispecific antibody targeting HER2 and T cells inhibits breast cancer growth via CD4 cells[J]. Nature, 2022, 603(7900): 328-334.
[13] Stadler C R, Bähr-Mahmud H, Celik L, et al. Elimination of large tumors in mice by mRNA-encoded bispecific antibodies[J]. Nature medicine, 2017, 23(7): 815-817.
[14] Lefèvre A, Parra-Guillen Z P, Trocóniz I F, et al. Mechanistic PKPD modeling to describe cytokine release associated with CD3 T-cell engager therapies[J]. Frontiers in Immunology, 2025, 15: 1463915.
[15] http://www.ema.europa.eu/en/medicines/human/EPAR/blincyto
[16] Zuch de Zafra C L, Fajardo F, Zhong W, et al. Targeting multiple myeloma with AMG 424, a novel anti-CD38/CD3 bispecific T-cell–recruiting antibody optimized for cytotoxicity and cytokine release[J]. Clinical Cancer Research, 2019, 25(13): 3921-3933.
[17] Trang V H, Zhang X, Yumul R C, et al. A coiled-coil masking domain for selective activation of therapeutic antibodies[J]. Nature biotechnology, 2019, 37(7): 761-765.














加入订阅
获取药物代谢与药代动力学最新专业内容和信息

